1.神经递质释放
NMJ 将运动轴突携带的电信号通过化学信号(神经递质 ACh)转换回肌纤维上的电信号,进而诱发肌肉收缩。这一过程依赖多种电压门控和配体门控离子通道,它们精确的功能特性和定位至关重要。NaV1.6 型电压门控钠通道将神经动作电位沿有髓运动轴突传导至神经末梢,由此引发的去极化激活活性区的部分 CaV2.1 型电压门控钙通道。内流的 Ca²⁺与 Ca²⁺激活蛋白结合,启动含 ACh 小泡的受控胞吐过程。这是一个复杂的重排过程,使小泡与突触前膜融合并将 ACh 释放到突触间隙,随后小泡回收并重新填充,该过程受多种蛋白调控。
通常,在极短暂的突触前动作电位期间,活性区仅约 20% 的 CaV2.1 通道会开放。每次神经动作电位释放的 ACh 小泡数量(“量子含量”)因哺乳动物物种和肌肉类型而异,大致在 20–100 之间。KV3.3 型电压门控钾通道的激活使神经末梢复极化,终止 ACh 的囊泡释放。当运动神经元被激活时,人类的发放频率为 20–40Hz,啮齿动物为 20–100Hz。在这些高强度放电过程中,ACh 释放量会下降 20–30%,这显然是由于神经胞吐机制中的速率限制步骤所致。
除了神经动作电位诱发的多个 ACh 量子同步释放外,还存在单个 ACh 量子的自发释放,这很可能是由于不受控制的溢出胞吐导致。小鼠 NMJ 处的自发释放速率约为 1 次 / 秒,部分依赖于 CaV2.1 通道的(自发)Ca²⁺内流。
2 ACh 受体与突触后电生理机制
释放的 ACh 扩散穿过突触间隙,与释放位点对面、突触后膜皱褶顶部聚集的肌肉 AChR 结合。肌肉 AChR 可以说是神经肌肉传递中最关键的成分,属于半胱氨酸环配体门控受体家族的异五聚体离子通道受体。事实上,正如 MG 和 AChR 缺陷型先天性肌无力综合征(CMS)中所见——亚基基因突变限制或阻止 AChR 表达,导致突触后膜 AChR 无法高度聚集——会引发易疲劳性肌无力。
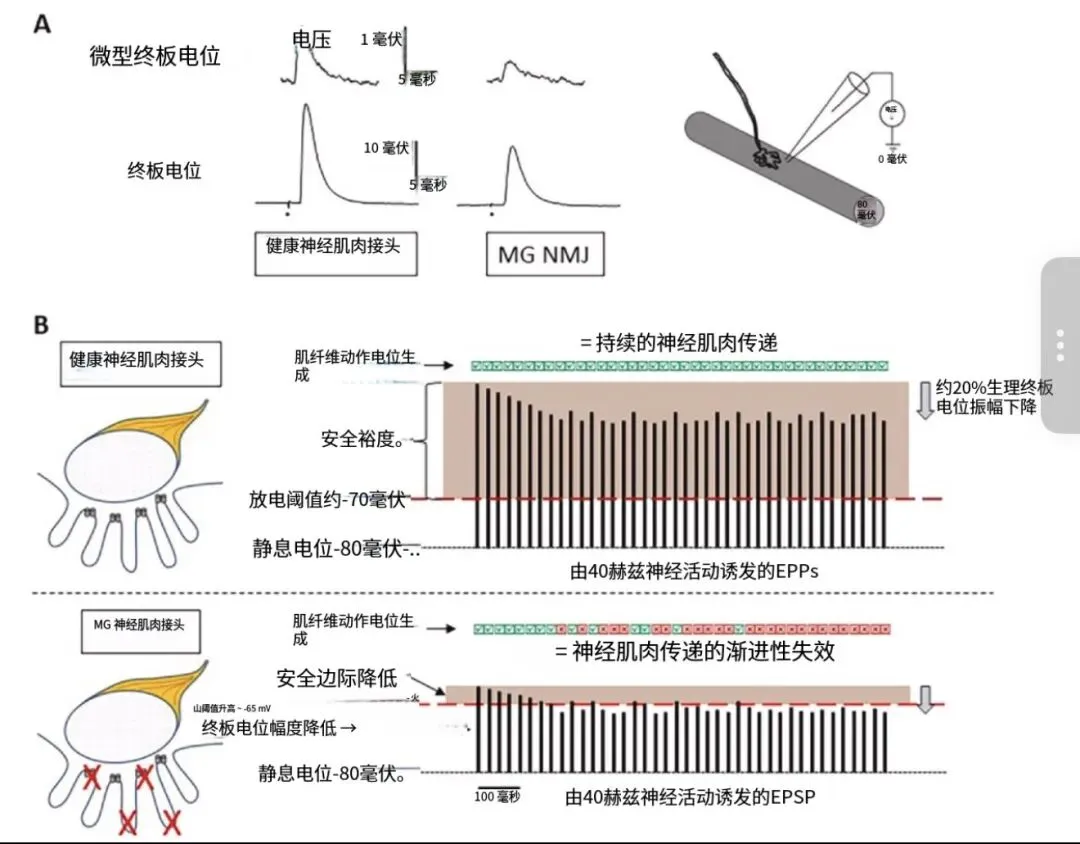
肌肉 AChR 存在两种同工型,其电生理特性不同,取决于五聚体中包含的是 γ 亚基还是 ε 亚基。含 γ 亚基的 AChR 调控发育中(和再生中的)NMJ 的神经肌肉传递,而含 ε 亚基的 AChR 则存在于成熟稳定的 NMJ 中。一个复杂的多步骤过程确保 AChR 的高密度聚集。肌肉 AChR 是一种变构膜蛋白,至少存在三种状态:关闭态、开放态(导电态)和脱敏态。两个 ACh 分子的结合可短暂稳定开放态,允许阳离子跨膜流动。Na⁺离子的主要内流产生约 20–30mV 的膜去极化,即 NMJ 突触后皱褶处的终板电位(EPP,图 2A)。
随后,AChR 激活的终止至关重要,这一过程通过突触间隙中乙酰胆碱酯酶对 ACh 的酶解实现。因此,EPP的持续时间在很大程度上取决于肌肉 AChR 的动力学特性。然而,ACh 释放增加或 ACh 降解减慢可通过扩大 ACh 的空间扩散范围,增加与 AChR 结合的 ACh 数量,最终增强膜去极化程度。EPP 精确的持续时间和幅值对传递系统的正常功能至关重要。事实上,AChR 亚基的突变会改变离子通道的动力学特性(CMS 的一种亚型),可能导致 EPP 缩短(快通道型 CMS)或去极化延长(慢通道型 CMS),两者均会导致神经肌肉传递的易损性。
EPP 产生后,会激活位于突触后皱褶深处的 NaV1.4 钠通道。其密度决定了发放阈值,约为 - 70mV(图 2B)。触发的肌纤维动作电位从 NMJ 向两侧沿肌膜扩散,激活胞内 Ca²⁺信号,通过肌动蛋白和肌球蛋白的相互作用引发肌纤维收缩。
突触前膜自发释放单个 ACh 量子,会导致有限数量的 AChR 开放,产生微小终板电位(MEPP,图 2A)—— 一种小幅去极化(约 0.3–1.5mV,因物种、肌肉类型、年龄等因素而异),其幅值低于阈值,不会触发动作电位。据目前所知,MEPP 无生理功能。
3 神经肌肉传递的安全系数
NMJ 的结构设计具有 “冗余性”,其产生的 EPP 远大于激活足够 NMJ 驻留 NaV1.4 钠通道以引发动作电位所需的去极化阈值。这种不平衡被称为安全系数,通过前文所述关键分子的精确定位和高密度分布实现。安全系数确保了神经肌肉传递的稳健性和可靠性,即使在高频放电时(ACh 释放减少导致 EPP 有所衰减)也能正常传递(图 2B)。如果安全系数降低(如 MG 中),神经肌肉传递可能失败,导致易疲劳性肌无力(图 2B)。肌肉内和个体间安全系数的幅值差异,可能是神经肌肉传递可靠性受干扰的易感性存在差异的基础。破坏 NMJ 成分结构、功能或定位的自身抗体,就是这类可将安全系数降低至临界水平的干扰因素。