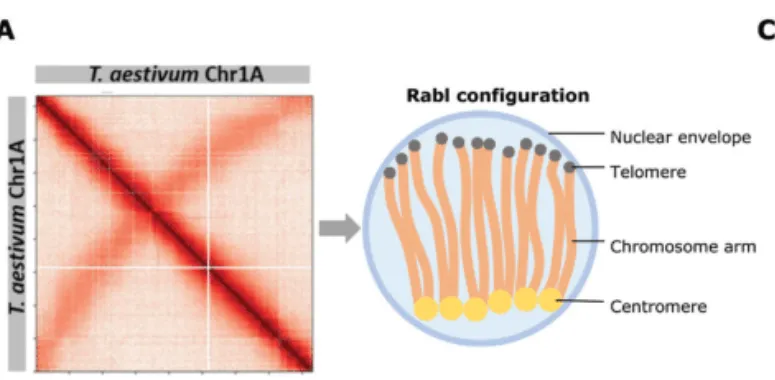
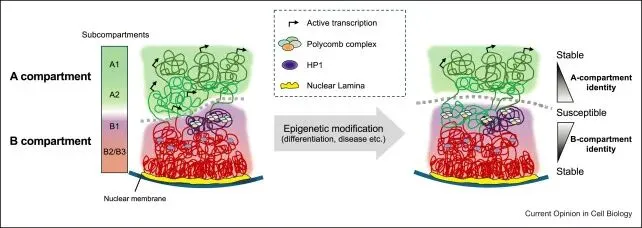
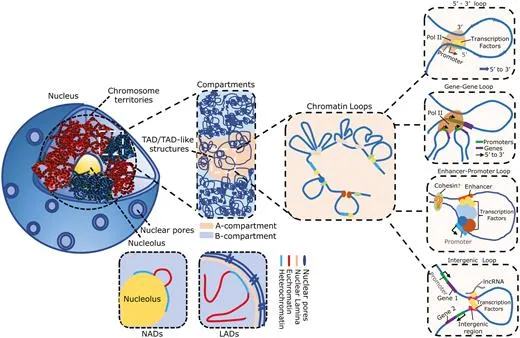
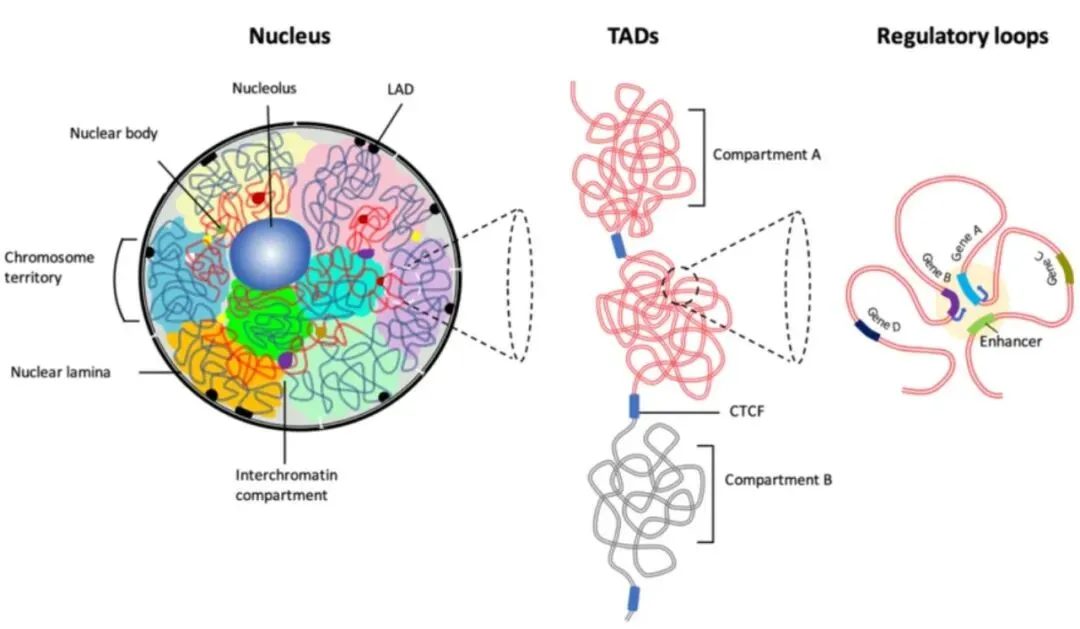
开头碎碎念:又又又又要学新东西而且时间很紧,和大家分享一下一些学习过程中需要理解记忆的基本概念吧。算是新开的一个part除了看的文献之外再做一些拓展,也希望能给大家带来一些帮助参考Rabl-like 构型(Rabl-like configuration):染色体在细胞核中呈现出着丝粒(centomere)聚集于核的一侧,而端粒(telomere)聚集于另一侧的极性排列方式Jet-like染色体互作( jet-like chromatin contacts):在Hi-C图谱中域对角线垂直的线状接触增强,这些结构代表了cohesin推动未受阻双向loop挤出过程,是loop extrusion在Hi-C数据中的动力学呈象特征。Chromosome Territories, CT(染色质领地):是指每一条染色体在细胞核内占据相对独立、非随机的空间区域,不同染色体之间的空间分布具有明显的分隔性,属于整个染色体尺度的空间组织方式,通过FISH等空间成像技术最早发现,为染色体内部更精细的三维结构提供整体架构。是当前认知中最高级最宏观的三维基因组组织形式。A/B compartment(A/B染色质区室):指在单条染色体内部,根据染色质的转录活性状态和空间互作偏好,将基因组划分为转录活跃的A区室和转录抑制的B区室,从而形成的大尺度空间分离结构,尺度通常为Mb级别,A区室倾向于彼此相互接触,B区室依然,常通过Hi-C数据的主成分分析(PC1)识别,与基因表达水平和组蛋白修饰状态高度相关。是染色体领地内部最主要的宏观分区方式之一。在高分辨率Hi-C数据中,A/B区室可进一步细分为多个亚型(如A1、A2、B1、B2等),反映不同程度的转录活性或抑制机制。核结构相关染色质区域(Nuclear structure–associated chromatin):此类宏观组织方式描述的是染色质在细胞核中相对于特定核结构的空间定位特征。核纤层相关结构域(Lamina-Associated Domains, LADs):指与核纤层(nuclear lamina)稳定结合的基因组区域,通常表现为染色质致密、转录活性较低。多与B区室重叠,富集抑制性组蛋白修饰(如H3K9me2/3),在基因表达调控中起到空间抑制作用
核仁相关结构域(Nucleolus-Associated Domains, NADs): 是指定位于核仁周围的染色质区域,通常富含重复序列和低表达基因。与转录抑制状态相关,在染色体空间定位中具有稳定性。
核斑点相关染色质(Speckle-associated chromatin):该类区域靠近富含转录和剪接因子的核斑点(nuclear speckles),通常与高转录活性区域相关。多属于 A 区室,与高表达基因密切相关。
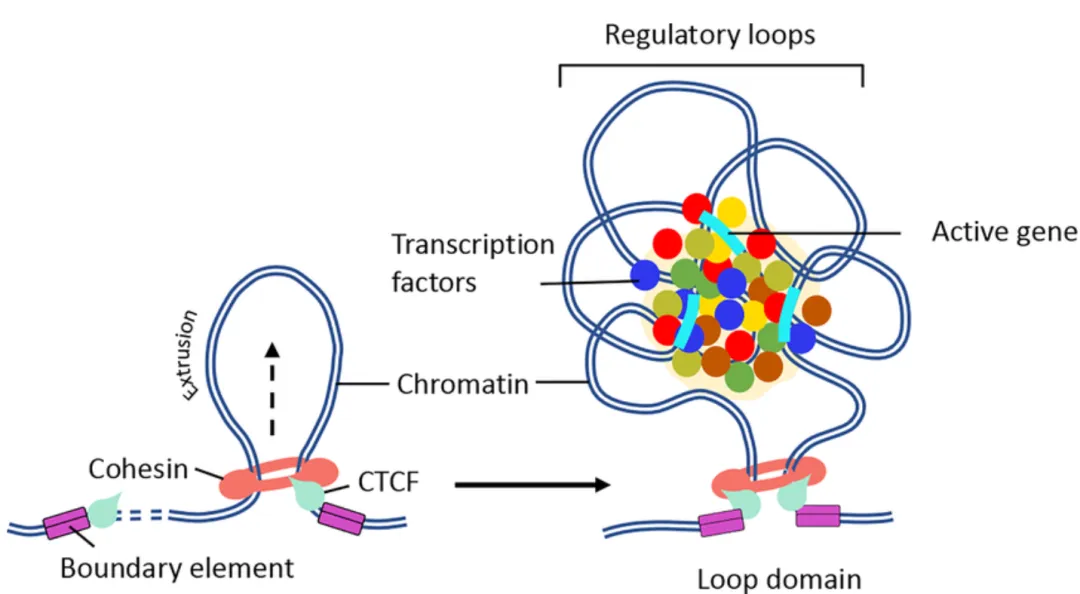
TAD(Topologically Associating Domain,拓扑关联结构域):TAD是染色体在三维空间中形成的中尺度结构单元,其特征是在该区域内部染色质片段之间的相互作用频率显著高于相邻区域的相互作用。是染色质的功能性调控单元。TAD可限制增强子作用范围,增强子通常只调控同一TAD内的基因,保证基因表达的特异性,TAD边界破坏可导致表达异常,TAD边界在不同细胞类型中相对保守,其内部活性状态(表达、histone)可动态变化。
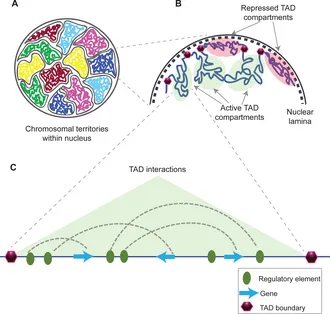
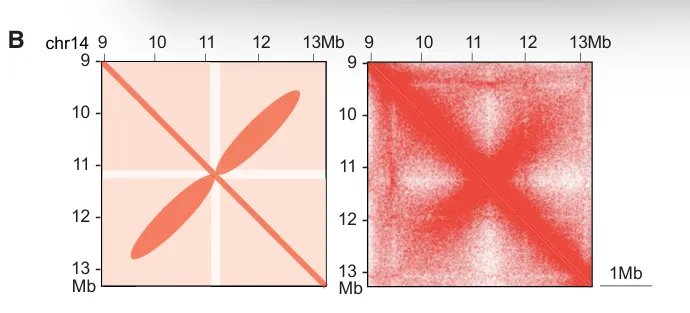
loop(染色质环,Chromtain loop):染色体上两个相距较远的基因组位点在三维空间中发生稳定或半稳定物理接触而形成的局部结构,常见类型有增强子-启动子loop,CTCF-CRCF loop,染色体结构loop,其可以直接调控基因转录,增强子域靶基因的物理基础,快速动态调控。一些结构之间的关联(针对The jet-like chromatin structure defines active secondary metabolism in fungi):jet-like domain(文章中给的新定义):在 Fusarium graminearum(镰刀菌)基因组中,活跃的次级代谢基因簇(BGCs)会形成一种在 Hi‑C 图谱上呈现 “V‑形”或类似喷射状(jet‑like)的局部交互结构域。这些结构域在 Hi‑C 图谱中呈对角线附近对称“喷射状”接触富集;可以向两侧对称延伸(propagate)数十 kb(最多约 54 kb);在全基因组范围内 普遍存在于活跃表达的基因区域;与 高水平基因转录、染色质可及性增强、组蛋白乙酰化标记富集 密切相关;是一种既定的、可识别的染色质局部交互域,而不是仅仅指动态的挤出过程;其形成依赖于组蛋白乙酰转移酶 GCN5 的存在,GCN5 缺失会阻断这些结构域的形成。Ouyang, W., Xiong, D., Li, G., & Li, X. (2020). Unraveling the 3D genome architecture in plants: present and future. Molecular Plant, 13(12), 1676-1693.Asami Oji, Linda Choubani, Hisashi Miura, Ichiro Hiratani. (2024). Structure and dynamics of nuclear A/B compartments and subcompartments, Current Opinion in Cell Biology,Volume 90,2024
Ying Huang, Natalia Yaneth Rodriguez-Granados, David Latrasse, Cecile Raynaud, Moussa Benhamed, Juan Sebastian Ramirez-Prado, The matrix revolutions: towards the decoding of the plant chromatin three-dimensional reality, Journal of Experimental Botany, Volume 71, Issue 17, 17 August 2020, Pages 5129–5147, https://doi.org/10.1093/jxb/eraa322
Guo et al., Chromatin jets define the properties of cohesin-driven in vivo loop extrusion, Molecular Cell (2022),https://doi.org/10.1016/j.molcel.2022.09.003
Matharu N, Ahituv N. Minor Loops in Major Folds: Enhancer-Promoter Looping, Chromatin Restructuring, and Their Association with Transcriptional Regulation and Disease. PLoS Genet. 2015 Dec 3;11(12):e1005640. doi: 10.1371/journal.pgen.1005640. PMID: 26632825; PMCID: PMC4669122.
Pongubala JMR, Murre C. Spatial Organization of Chromatin: Transcriptional Control of Adaptive Immune Cell Development. Front Immunol. 2021 Mar 29;12:633825. doi: 10.3389/fimmu.2021.633825. PMID: 33854505; PMCID: PMC8039525.
Wenyong Shao, Jingrui Wang, Yueqi Zhang, Chaofan Zhang, Jie Chen, Yun Chen, Zhangjun Fei, Zhonghua Ma, Xuepeng Sun, Chen Jiao, The jet-like chromatin structure defines active secondary metabolism in fungi, Nucleic Acids Research, Volume 52, Issue 9, 22 May 2024, Pages 4906–4921, https://doi.org/10.1093/nar/gkae131